
转基因油菜籽(Brassica Napus L.)中CAMV 35s启动子表达模式的组织化学分析
Parissa Jonoubi.1*,Ali Hatef Salmanian2和amin ravaei.1
1伊朗德黑兰生物科学学院植物生物学系,生物科学学院。
2植物生物技术,国家基因工程研究所和生物技术研究所(Nigeb),德黑兰,伊朗。
http://dx.doi.org/10.12944/CWE.10.Special-Issue1.90
促进剂被认为是生物技术的有价值的工具,为优雅目的提供了巨大的机会。本研究旨在探讨CAMV 35S启动子在转基因油菜籽中的GUS基因的表达模式(芸苔栗鸟l .)。将GUS基因转移到油菜外植体上农杆菌.再生的和有根的转基因植株被转移到含有泥土和蛭石混合物的花盆中。对这些植物进行了PCR和组织化学GUS检测。获得了转化植株的叶、叶柄和茎的横截面。GUS在韧皮部、薄壁组织、厚角组织和维管束支持组织中均有活性。形成层、内胚层、维管射线、髓薄壁组织、表皮和毛状体中也有。这些结果表明,CaMV 35S对转基因在不同组织中的表达有不同的影响。
维管束;GUS;组织培养;转型;农杆菌
复制以下内容以引用本文:
Jonoubi P,Salmanian A.H,Ravaei A.转基因油菜籽(Brassica Napus L.)中Camv 35s启动子表达模式的组织化学分析。Curr World Environ2015的特刊; 10(2015年5月特别问题)。DOI:http://dx.doi.org/10.12944/CWE.10.Special-Issue1.90
复制以下内容以引用此URL:
Jonoubi P,Salmanian A.H,Ravaei A.转基因油菜籽(Brassica Napus L.)中Camv 35s启动子表达模式的组织化学分析。Curr World Environ2015的特刊; 10(2015年5月特别问题)。可从://www.a-i-l-s-a.com?p=680/
介绍
由于植物的优点,包括高科核系统对植物系统的外来基因表达和重组蛋白质产生的影响,包括高产品质量,成本非常低,扩大量高,糖基化的轻微差异,低风险污染,相对较低的道德问题,廉价的储存成本[(Ma等,2003) - (Egelkrout等,2012)]。根据国际服务的国际服务,收购Agri-Biotech申请(ISAAA)报告“2012年,从1996年的170万公顷的170万公顷达到了前所未有的100倍,2012年的170万公顷 - 这使得生物技术作物最快的作物技术在近期历史 - 原因 - 他们提供福利“(James等,2012)。为了转基因植物,需要调节基因上游的调节启动子以确定外国基因表达的部位,水平和时序(Egelkrout等,2012);这些启动子通常分为五组:1)组成型启动子,其在大约所有细胞类型中导致最高量的表达。在大多数植物细胞中高度表达的植物生物技术中最广泛应用的组成型启动子是花椰菜马赛克病毒(CAMV)35s启动子(Yoshida等,2000)。2)诱导型启动子,其导致在存在或不存在因子中的存在或不存在诸如伤害或病原体侵袭,化学或物理诱导的因素中激活或停用,并且优先于没有干扰转基因的生命周期的发育阶段植物(Corrado等,2009)。3,4)组织特异性和显影 - 阶段特异性启动子是将转基因在特定组织中的表达引起的那些,例如叶片,种子,根部等或在植物发育的特定阶段,在某些情况下, increase the stability of the produced protein [(Ma et al., 2003)- (Egelkrout et al., 2012)]. 5) Synthetic promoters which are composed of the regulatory elements of natural promoters (Sharma et al., 2009).
重组蛋白的定位是植物转化的相当方面,可能影响蛋白质的保存和纯化程序(Wilken等,2012)。考虑到所选择的启动子,外来蛋白质的定位将是可预测的,并且报告基因的应用将促进该程序。使用GUS基因作为报告者,可以评估新型启动子的功能,以便可以对基因表达的急性控制进行适当的选择。此外,常规组织化学分析的可用性将极大地促进植物发育过程中基因表达模式的研究。此外,这些方法能够更快且敏感地筛选转化的细胞和组织。
作为组成型启动子,CAMV 35S启动子应该自然地将外源基因的表达直接引导到大约所有细胞和组织类型中;然而,存在一些证据表明,特别是植物的特定细胞和组织的不同基因表达意味着不同的表达模式(Anuar等,2011)。
本研究旨在检测转基因油菜不同组织中GUS的活性,以揭示CaMV 35S启动子定向基因的表达。
材料和方法
这农杆菌肿瘤术含有PBI121二元载体的菌株LBA4404用于植物转化。载体的T-DNA区域有NPT IICaMV 35S启动子控制下的GUS基因。
转型子叶植物的外植体农杆菌
这芸苔栗鸟var。PF 7045-91和SLM种子在表面灭菌后,在含有半强度MS的罐中发芽(Murashige等,1962)培养基,并在40-50μm的16/8h光周期中保持-2S.-1光照强度和25°C的温室。这农杆菌在LB (Luria-Bertani)肉汤中(含50mg-1在28℃下摇晃(180rpm)的卡那霉素。这农杆菌文化与OD650.= 0.8以3000rpm离心15分钟。将颗粒重悬于感染培养基含量,在pH5.2处的5%葡萄糖,并用于转化外植体。仔细兴奋,子叶(带5-7毫米的叶柄)7天龄幼苗和14天幼苗的幼苗。只接种子叶子和幼杆块的叶柄农杆菌悬挂5至10秒。将所有外植体用滤纸擦拭干净,置于pH 5.2不含任何激素的MS培养基中,置于温室,28℃,暗条件下保存。共培养后,将下胚轴外植体转移到愈伤组织诱导培养基(CIM) (MS培养基含1mg-12,4 - d, 30 gl-1蔗糖,7原-1琼脂,200毫克-1头孢噻肟和15米格兰-1Kanamycin在pH5.8)中,然后转移到芽诱导培养基(SIM)(MS培养基含有1.5 mgL-1苄基腺嘌呤,30 gl-1蔗糖,7原-1琼脂,200毫克-1头孢噻肟和15米格兰-1Kanamycin在pH5.8),并在相同的新媒体上每两周转移一次。在共培养后直接将子叶蛋白质外植体转移到SIM培养基中。6周后,从子叶植物的外植体激发绿色卡霉素抗芽并转移到芽成熟培养基(SMM)(含有MS盐的MS培养基,20GL-1蔗糖,7原-1琼脂、15球型-1卡那霉素,200 mg-1头孢噻肟pH为5.8)。2周后,将绿芽转移到根诱导培养基(RIM) (MS + 2mgl)中-1吲哚丁酸,20 gl-1蔗糖,6 GL-1琼脂,200毫克-1头孢噻肟和15米格兰-1Kanamycin在ph 5.8)。在开发根部后,能够产生根的卡那霉素抗性绿植物分别转移到蛭石,蛭石和土壤混合物和土壤中。
转基因植物GUS基因的PCR验证
根据CTAB方法(Murray等,1980)萃取卡那霉素抵抗植物的基因组DNA,以便用于转基因植物中的GUS基因。利用包括GUS + 2和GUS -4的特异性引物。引物序列如下:
GUS-4: 5“-CCGGCATAGTTAAAGAAATCATG-3”
格斯+ 2:5“-GGTGGTCAGTCCCTTATGTTACG-3”
用TECHNE termocycler进行扩增:94℃变性1 min, 61℃退火1 min,引物72℃延伸1 min, 72℃终延伸7 min, 94℃初始变性4 min,共30个循环。
GUS测定
在含有50 mM磷酸盐缓冲液(pH 7)、1 mM X-Gluc、1 mM EDTA、0.001% triton和10 mM β-巯基乙醇的缓冲液中,对GUS基因表达和β-葡萄糖苷酶(X-Gluc分解产生蓝色)的产量进行了评价。转基因植株的茎、叶柄和叶片在37℃的缓冲液中孵育过夜。为了观察植物组织中的蓝色,用96%乙醇去除叶绿素。
转化分析
通过手动横截面分析转基因植物的营养组织。将所得部分通过胭脂碱 - 背心染色并使用光学显微镜观察。
结果与讨论
使用农杆菌-介导转化,得到PF7045-91和SLM品种的转化植株,转化率分别为10.2%和12%。图1显示了转化和未转化的植物。未转化植株在选择培养基上变成白色/紫色;转化植株保持绿色,表型正常。
 |
图1:Kanamycin作为子叶植物外植体的选择剂的影响a)转化的植物在卡那霉素存在下保持绿色;b)在卡那霉素存在下,未转化的植物在白杨中成为白色/紫。 点击此处查看数字 |
对GUS基因的PCR验证表明,该基因存在于所有假定的转基因株系上。图2为显示GUS基因的520 bp PCR产物。这一结果表明,二元载体的T-DNA区域已成功转移到植物细胞中。
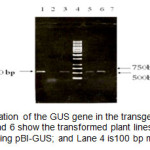 |
图2:GUS基因在转基因植物中的PCR扩增。第3、5和6道是经过改造的植物线;Lane 1是pBI-GUS阳性对照;4号道是100 bp的混合梯。 点击此处查看数字 |
转基因植株的组织呈现蓝色,表明GUS基因的表达和β-葡萄糖醛酸酶的功能。在73%的抗卡那霉素植株中,在不同强度下均呈现蓝色(图3)。在叶片横截面上,维管束中GUS酶活性极高,除韧皮部外,维管束射线和髓薄壁组织均呈现极显著的蓝色。皮层和髓的实质细胞以及表皮细胞(包括毛状体)的染色也不同。
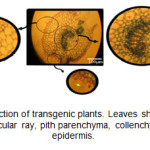 |
图3:转基因植物的叶横截面。叶子展示GUS表达在血管束,血管射线,髓薄膜,Collenchyma,richomes和表皮。 点击此处查看数字 |
在转基因植物的叶柄横截面中,在Phloem和Collenchyma组织中观察到蓝色(图4)。
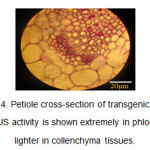 |
图4:转基因植物的叶柄横截面。GUS活动在Collenchyma组织中非常庞大显示。 点击此处查看数字 |
在茎横截面中,在Phloem组织和血管射线中高度观察到由X-Gluc分解产生的GUS基因和蓝色的外观(图5)。Gus活性在Camv-Gus植物杆的血管环周围的Phloem组织中高度看出(Jefferson等,1987)。表达模式在接合内部和外部验证的Phloem薄壁症中标注和观察到(esauk等,1977)。在一些转化植物的茎中,观察到GUS活性的可变分布。
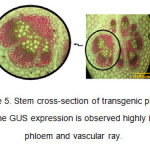 |
图5:转基因植物的茎横截面。在韧皮肌和血管射线中高度观察到GUS表达。 点击此处查看数字 |
绝对清楚的是,GUS表达的模式在植物细胞中复杂(Basu等,2004)。预计在不同细胞类型的植物中预期观察到不同的代谢活动;因此,GUS基因表达受不同的生化,分子和生物因子(Fior等,2009)的影响,以转录和翻译的速率展出;目前的发现可能暗示这种差异。或者,由于Phloem细胞的非常小的横截面积,观察到的强烈的蓝色可以指每个单位区域的更大细胞数(Jefferson等,1987)。在细胞周期的S期间,CAMV 35s启动子如此优选地活跃,使得与观察到的GUS表达模式的解释可能意味着这些组织内的细胞分裂(Nagata等,1987)。本备注来自于在病毒复制中的CAMV的35S脚本作用;一些其他植物DNA病毒如,Geminiviruses在韧皮组织中具有如此复制[(Schubert等,2004)和(Dinant等,2004)]。此外,Camv 35s启动子具有两个结构域:结构域A有助于共同组织表达和结构域B导致血管束表达(Anuar等,2011)。考虑到较高植物中的低或没有内源性GUS活性,观察到的定位可以反映不同细胞中CAMV 35s启动子的表达水平的实际差异。
使用农杆菌介导的转化,具有不同副拷贝的转基因细胞和外源基因的植物以及不同的整合位点。因此,转化的植物将表达具有不同量的外源基因。这种差异可能是由于基因集成的各种定位,内源性蛋白因子的影响,以及在转录或转录过程中沉默[(Wakimoto等,1998),19和(Schubert等,2004)]。高稳定性基因插入率可能导致基因表达水平不稳定或降低。转型农杆菌将导致低速率的外源基因插入和DNA积分将随机随机,其倾向于在基因组的远端染色体区域中插入[(Dong等,2001)和(Gelvin,2003)]。
现在,人们对转基因植物及其产生外源蛋白的特性有了深刻的认识。如果没有合适的启动子,外源基因将被复制,但不会表达;因此,使用合适的启动子将导致基因的成功表达。尽管CaMV 35S启动子是一个强大的启动子,但考虑到基于细胞类型和细胞周期的转录过程,它不应该被认为是“组成型”的。大量启动子的可用性使得利用植物作为生产外源蛋白质的工厂的前景非常广阔。目前,关于植物转化和外源基因表达的研究进展良好,今后的工作可能会更有启蒙性。
致谢
我们感谢Kharazmi大学植物发育生物学实验室和NIGEB植物生物技术实验室的工作人员提供的所有援助。这项研究得到了Kharazmi大学和国家遗传工程和生物技术研究所(NIGEB)的支持。
参考文献
- Ma K.J. Drake M.P. Christou P., Nat Rev. Genet。, 4794(2003)。
- 沙玛·a·k和沙玛·m·k,生物技术公司。中国海洋大学学报(自然科学版),2014,34(1):1 - 8。
- 埃格克劳·拉詹·霍华德,《植物》。科学。, 184,83(2012)。
- James C., ISAAA Brief,(2012)。
- Yoshida K. Shinmyo A.,J.Biosci。生物。,90,353(2000)。
- Corrado G. Karali M,生物技术。地理科学进展,27,733(2009)。
- 转基因植物系统的下游加工威尔康L. Nikolov Z.蛋白质回收和净化策略。植物中的分子养殖:最近的进展和未来的前景,斯普林斯,荷兰,(2012)。
- 王志刚,王志刚,王志刚。, 10,8236(2011)。
- Murashige T. Skoog F.,Physiol。Plantarum。,15,473(1962)。
- 默里米。汤普森W.F.,核。酸。Res。,8,4321(1980)。
- 杰斐逊r.a.kavanagh t.a.Bevan M.W.,Embo。J.,6,3901(1987)。
- 《种子植物的解剖》,《植物学报》,1998年第1期。
- Basu C. Kausch A.P.Chandlee J.M.,Biochem。Biophys。res。公共交流。,320,7(2004)。
- Fior S. Gerola P.D.,植物。方法,5,1(2009)。
- Nagata T. Okada K. Kawazu T. Takebe I.,MGG。,MGG。,207,242(1987)。
- 舒伯特D. Lechtenberg B. Forsbach A. Gils M. Bahadur S. Schmidt R. Plant。细胞。, 16,2561(2004)。
- 迪南特·里波尔·皮珀,《生理学》。杆菌。, 121,108(2004)。
- Wakimoto B.T。细胞。, 93,321(1998)。
- 汪格E.J.加西亚-布兰科m.a.,《细胞》杂志。医学杂志。, 213281(2001)。
- 董建杰哈伯平邓文敏霍尔t.c.,摩尔育种。, 7,187(2001)。
- Gelvin S.B.,MMBR。,67,16(2003)。

这项工作是在授权下获得的知识共享署名4.0国际许可.





