
电磁波对玉米光合色素和抗氧化酶的影响
Habibeh Zare1和卅三申请的请求项1*
1生物系,设拉子大学,伊朗。
http://dx.doi.org/10.12944/CWE.10.Special-Issue1.88
为了研究Zea Mays L中的抗氧化光合色素含量的电磁波长对抗氧化光合色素含量的影响,每8小时通过电磁波(200 Gus强度)照射含水5小时5小时的含有湿的种子的培养皿。每8小时大约半小时。治疗后,将种子转移到Perlit和Pitmass中。研究了13天的叶子。与对照相比,叶片引起的丙酮提取物的化学分析表现出下处理样品的叶绿素A和B的量减少。对照和湿法处理的差异有意义,但对照和干处理样品之间没有任何显着差异。在处理样品下显示出与对照样品相比,类胡萝卜素和非酶促抗氧化剂如酚类化合物,黄酮类化合物和脯氨酸的量增加。此外,在酶促抗氧化剂如过氧化氢酶,抗坏血酸过氧化物酶和与湿法处理样品相关的比较控制的情况下,观察到显着增加。然而,OFF系统治疗样品和对照之间没有显着差异。
电磁波;非酶抗氧化剂;酶抗氧化剂;光合色素;Zea Mays L.
复制以下内容以引用本文:
Zare H.和Mohsenzadeh S.电磁波对玉米光合色素和抗氧化酶的影响。DOI:http://dx.doi.org/10.12944/CWE.10.Special-Issue1.88
复制以下内容以引用此URL:
Zare H.和Mohsenzadeh S.电磁波对玉米光合色素和抗氧化酶的影响。可从://www.a-i-l-s-a.com/?p=9983.
介绍
生物和非生物胁迫通过诱导氧化应激不同程度地影响植物的代谢(Apel和Hirt, 2004)。电磁波传递能量,所以它们可能会影响包括植物在内的生物体。研究表明,电磁波对植物生命的各个方面都有影响,如植物的发育和生长、繁殖和细胞结构的功能等。根据辐射的频率和转移到植物细胞的能量的多少,植物的反应是不同的。低频电磁波改变种子的生存能力而不造成任何硬损伤(Yao et al., 2006)。像玉米这样的植物通常不是从种子中生长出来的,它们的增殖是通过根的分裂、茎的繁殖和芽的生长来完成的。因此,应准备适宜的发芽条件。引种是一种促进种子萌发的方法,在逆境条件下也能促进种子萌发。显然,在这种方法中,包括酶激活在内的萌发的第一个阶段是通过浇水的种子通过的,当然没有任何细根。此外,电磁波具有诱导活性,可以通过影响细胞核基因、促进代谢、增加酶活性和水分吸收来加速萌发
电磁波还作为胁迫诱导剂,通过氧化胁迫改变受胁迫植物细胞的化学含量,增加活性氧(ROS)。植物产生不同的抗氧化剂化合物并将其储存起来,以缓解压力,增强抵抗力。类胡萝卜素含量的增加是逆境胁迫下植物的一种防御反应。类胡萝卜素被认为是植物光合作用和叶绿体设备的保护者(Tevini et al., 1991)。胁迫下类胡萝卜素生物合成的增加可能会减少波浪对叶绿素色素的破坏效应(IIao,1997)。为了提高防御机制,植物在其组织中产生和聚集酚类物质,并储存具有不同结构和生化性质的黄酮类化合物,利用多种机制清除自由基(Shao et al.,2008)。脯氨酸的产生也是逆境植物的另一个重要适应。脯氨酸具有不同的清除细胞ROS的机制。脯氨酸在高等植物中的聚集与活性氧的增强直接相关(Maggio et al., 2002)。胁迫下植物的另一个防御机制是增加酶抗氧化剂的活性,如过氧化氢酶、抗坏血酸过氧化物酶和超氧化物歧化酶。 Prostetic groups of these enzymes have an important role in scavenging free radicals(Alscher et al.,2002). As Zea mays L includes effective chemical compounds, the purpose of this study is to determine if stress of low intense electromagnetic waves could result in enhancement of plant without plant destructing.
材料和方法
本研究选用Zare研究所524杂交种所提供的玉米种子。灭菌后的培养皿包括种子放在H2O2溶液中作为渗透素体。制备渗透素溶液为100ml蒸馏水。将种子置于渗透溶液中5h,启动后弃去渗透溶液,用蒸馏水洗涤种子3次,用滤纸取剩余水分,实验室温度烘干,为下一步准备。部分种子在100 ml蒸馏水中浸泡5h。在本研究中,包括湿种子在内的培养皿水平放置在两个磁片的表面,在泰万的电磁波工业系统中,每天和每次20 mT电磁波辐射30分钟。然后在pH=6和25的温度下,将种子转移到花盆中。测定了植物13 d叶片的光合色素、酚类化合物、黄酮类化合物、脯氨酸等非酶抗氧化剂的含量和过氧化氢酶、抗坏血酸过氧化物酶、超氧化物歧化酶等酶抗氧化剂的活性。采用Spss 16统计软件进行邓肯检验,4个重复进行处理与对照比较,p值采用Excel绘制图。
光合色素的测定:用Arnon法和Lichtenthaler法测定叶片中叶绿素a、b和类胡萝卜素丙酮提取物的含量。用5ml 80%丙酮将0.2克新鲜叶片组织完全磨碎,然后用Wattman滤纸过滤得到的溶液。然后再加入5ml丙酮,用分光光度计(SPEKOL 1500)在663、646和480 nm处测量得到的溶液的吸光度。光合色素和类胡萝卜素的浓度用下列公式测定。
的酚含量测定:麦当劳et.al的方法被用来测量酚含量。用5ml 1N福林 - siocalto0.5毫升甲醇提取物进行混合,然后碳酸盐4毫升钠1M溶液,加入蒸馏水,使之100cc的。溶液放置在黑暗中15分钟,并在765 nm处测定吸光度。没食子酸被用来描绘标准校准曲线。
黄酮类物质含量的测定:甲醇提取物的评估要考虑黄酮含量常法的基础上完成的。的甲醇提取物溶液0.5毫升由1.5cc甲醇的甲醇混合95%,氯化铝0.1毫升10%,0.1cc乙酸钾1M和2.8毫升去离子蒸馏水。30分钟后,吸光度415nm处进行测定。没食子酸被用来描绘标准校准曲线。
脯氨酸含量测定:脯氨酸含量的测定通过Bates等人,方法完成。在该方法中0.5gr新鲜叶用sulfusalcilic酸10ml的3%溶液笑了。2毫升从均匀的混合物收集在过滤和除2毫升ninhydryn指示器加上2毫升大蒜愈伤酸后,封闭管被放置在双重蒸锅用100 tempretaure 1小时。1小时后,将试管置于冰上来停止反应。然后,4毫升甲苯加入到每个管中,并强烈搅拌。甲苯相的吸光度在520nm处测定,并由于它的标准曲线实现脯氨酸量。
过氧化氢酶活性的测量:Dhindasa的基础上已完成使用还原在H 2 O 2为240nm的吸光度估计过氧化氢酶的活性蚂蚁Motowe方法。该混合物包括的50mM磷酸钾pH值= 7和15mM的H2O2.then的30μL的2.87毫升,加入100毫升酶法提取开始反应。在吸收差异进行测定。在混合物的H 2 O 2量,使用猝灭常数ε= 0.28毫摩尔计算且A =εbc方程示出酶活性的程度。A代表吸光度,ε为淬火常数,c为浓度H2O2和b为妄想长度(1cm)为。酶活性为存在蛋白质(毫克)提取物每100μl的在1分钟内的总量来测量。
抗坏血酸过氧化物酶活性的测量:基于NAKANO和ASADE方法进行过氧化物酶活性的测量。混合物反应包括具有pH = 7,0.15m H 2 O 2,0.5mM抗坏血酸和50μL酶提取的50mM磷酸盐缓冲液。在抗坏血酸氧化与酶促反应开始后测量290mM的吸光度降低。常数被引入等于2.8米,并根据每分钟吸收变化的=εbc方程。
超氧化物歧化酶酶活性的测量:用于评估超氧化物歧化酶活性的Giancopitis和RIC提供的方法。反应混合物包括2.5ml 50mM磷酸钾缓冲液,pH = 7.8,0.1ml 13mM甲硫氨酸,0.1ml 75mm硝基丁基唑唑鎓,0.1ml2μm和0.2ml酶提取。将样品暴露于光线15分钟,并在560mM中测量它们的吸光度。在这项工作中,提供了两种控制样品,在不接收光的情况下提供的第一控制样品用作坯料的光和第二样品暴露于光的坯料15分钟,其由于酶存在而在光线下完全完成。超氧化物歧化酶活性的单位解释了一定量的酶,这导致硝基蓝四唑鎓的轻微还原抑制的50%。
结果
对照和处理样品中光合色素的测定说明了处理样品中叶绿素色素(a和b)数量的减少和类胡萝卜素含量的增加(图1)。根据统计考虑的结果,叶绿素a、湿处理B较对照分别下降7.45和18.35(图2),具有一定意义。off系统处理中叶绿素a、b含量下降分别为2.69和4.85,没有意义(图3)。比较湿处理和干处理样品中类胡萝卜素含量,有意义增加。湿处理比对照增加18.13,干处理比对照增加11.18。
根据文献记录的结果,与对照样品相比,干处理样品的酚类含量增加了85%,类黄酮含量增加了52.73%。干处理对酚类和类黄酮类化合物的影响分别为29%和31.42%。与对照组相比,两组治疗有显著差异(图4)。统计调查显示
如图5所示,在受胁迫植物的电磁波导致抗氧化酶活性的增加。这种增加在相对于对照湿处理过氧化氢酶,抗坏血酸过氧化物酶和超氧化物歧化酶酶计算55 39.80 24.11和分别是有意义的。相同的比较在干燥处理和控制其显示11.89,9.85和7.14增加过氧化氢酶,抗坏血酸过氧化物酶和超氧化物分别氧化物歧化酶,但差异不是有意义之间完成。
 |
图1:设备生产电磁波和它的工作方式 点击这里查看图 |
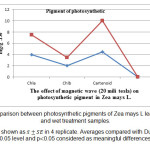 |
图2:ZEA的光合色素与控制和湿法处理样品的光合色素的比较。 结果显示为在4次复制中。平均值与0.05级和p的邓肯测试相比0.05被认为是有意义的差异。 |
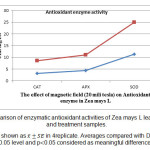 |
图3:的控制和处理样品在玉蜀黍叶子大号的酶促抗氧化活性的比较。 结果显示为在4张。平均值与0.05级和p的邓肯测试相比0.05被认为是有意义的差异。 点击这里查看图 |
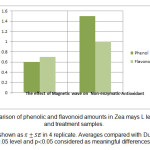 |
图4:玉米L叶中酚类和黄酮含量的比较。 点击这里查看图 |
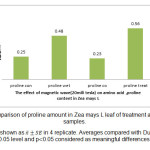 |
图5:玉米L叶脯氨酸处理与对照样品的比较 点击这里查看图 |
讨论
植物能探测到压力并作出反应。胁迫如电磁波会诱导植物产生抗氧化物质和改变植物的化学成分。这些化合物的增加表明植物细胞的活性是为了中和或调节逆境的影响,其结果是提高植物对逆境的抵抗力。植物和其他生物一样会对压力做出反应。根据Hosseinio和他的同事的理论,植物通过产生有效的抗氧化化合物来抵抗不同的应激因素。植物控制自由基最重要的防御机制之一是诱导酶促和非酶促抗氧化剂。研究表明,植物的许多代谢过程会产生ROS或自由基,但也存在有效的清除机制,如抗氧化机制(Blokina et al.,2003)。在正常情况下,活性氧产生量和抗氧化防御清除能力之间存在平衡。相反,在应激条件下自由基的产生量大于其清除能力。总的来说,抗氧化防御能力对这些因素的变化似乎是至关重要的(Smirnoff, 1995)。
在该研究中,观察到叶绿素A,B含量和增加应力植物的类胡萝卜素含量的降低。根据Jansen和Coworker的理论,由于抑制相关或通过在电磁波的应力下抑制叶绿素前体的抑制,可能发生叶绿素的降低。Caldwell和Coworkers还报告说,叶绿素还原的原因是叶绿素的降解或对叶绿素合成前体的负面影响。使用叶绿体吸收大量能量,光合膜很容易受损。如果能量未照相捕获,则会有保护机制。保护机构可用作传送额外能量的阀门。如果叶绿素的这种激发情况不会迅速抑制,因此可能与分子氧的可能反应,这导致激发的氧气(单线氧)(IIAO,1997)。叶绿体的类胡萝卜素可以保护这种细胞器免受波浪的破坏效应。这些化合物能够通过接受短波长的高能量并用作抗氧化剂来使ros失活。类胡萝卜素通过快速抑制兴奋的叶绿素来解决。 Excited carotenoids are not capable to form singlet oxygen and returns to initial state by losing energy as heat(Yao et al., 2006). Increasing of non-enzymatic antioxidant including phenol, flavonoid and proline are some kind of defensive mechanisms which happens in plants with stresses of electromagnetic waves (consistent with our results). On the basis of Caldwell and coworker's opinion, electromagnetic waves cause induction of cinamic acid synthesis and induce phenilpropanoid pathway that provides effective antioxidantresistance against stress in higher plants. Limitation in transferring of photosynthetic electron of stress condition seems to be one reason for flavonoids synthesis induction. Inhibition of flavonoid biosynthesis pathway using inhibitors of phenyl alanine amunialyase synthesis lead to increased sensitivity of plants to waves. In accordance with Tevini and coworker's study, flavonoids are important antioxidant compounds which neutralize free radicals and also prevents from their extra production. Production and aggregation of flavonoids in vacuoles of stem and leaf epidermal cells cause wave absorbance and protects organelles like chloroplast against waves which reduces oxidative stress effects(Flint et al.,1985).Sakihama et.al reported that high aggregation of phenolic compounds inside epidermal cells and their cell wall, also fuzzes and vacuoles of epidermal cells prevents from damage in inner mesophilic cells and is used for photosynthetic carbon assimilation with improving defensive mechanisms. Sakihama et.al using mutants unable to produce phenolic compounds revealed importance of these compounds in rice against electromagnetic waves radiation. Proline also as an effective defensive compound protects cells against stress of waves and in stressed plants increases as a defensive or adaptation response (Maggio et al., 2002). Proline plays different roles in water balance, stability of proteins, enzymes and their 3D structure. Proline also is carbon and nitrogen source for growing after removing the stress. This compound cause reduction in ROS threat, scavenging of free hydroxyl radicals, quenching of singlet oxygen, adjustment of cell pH and adjustment of /NADPH. Proline has a role in stability of membrane by integrating with phospholoipid membrane and changing hydrated layer around biological molecules. This substance is also effective in decreasing of thylakoid membranes damage by scavenging and reduction of free radicals (Verbruggen and Hermans 2008). Proline as a storage source of carbon and nitrogen the resistance of plants against stress plays an important role in adaptation and protective responses of plants(Siripornadulsil et al., 2002). Investigations also explain a direct relation between proline aggregation in higher plants and ROS increasing. This increase also may be the reason for proline production from glutamic acid(Hare and Cress,1997). Increasing of catalase, ascorbate peroxidase and superoxide dismutase activity as antioxidant enzymes, consistent with present study, is another defensive mechanism which happens in plants under stress of electromagnetic waves. Alscher et.al reported that prostatic group of homoprotein enzymes has a key role in removing of plant free radicals. Superoxide as a main ROS and a poison compound in cells cause enzyme nature variation, lipid oxidation and DNA disruption. Superoxide dismutase as a metalo-enzyme changes superoxide ion into hydrogen peroxide and molecular oxygen which reduces its destructive effects. This enzyme act as antioxidant defense line against free radicals but produces hydrogen peroxide that is another poison element(Alscher et al., 2002). Resulted hydrogen peroxide under superoxide dismutase function and a lot of other natural mechanisms of cell by enzymes such as catalase, ascorbate peroxidase are controlled. Catalase using direct impact of these compounds, makes hydrogen peroxide a substrate and prevents from its destructive effects by changing this molecule to water and molecular oxygen(Hare and Cress,1997). peroxidase are also a great family of defensive enzymes which catalyzes redox reaction between hydrogen peroxide as an electron acceptor and a lot of substrates such as phenolic compounds, ascorbat acid, aromatic amines and cytochrome c. plant has been attacked by free radicals less when this enzyme is increased(jonsen et al., 1998). In stressed plants, ascorbate peroxidase is increased in cytosol, vacuole, chloroplasts and apoplastas which plays an important role in balancing of produced free radicals amount(Zao and Chang,2008). In this compound, ascorbate is an electron acceptor and reducer element which plays an important role in scavenging of hydrogen peroxide(Gill and Tuteja, 2010).
结论
Zea Mays L种子的灌注提供了它们的发芽和电磁波的辐射导致萌发率增加。在ZEA中,可以对具有低强度的电磁波的反应,增加非酶促抗氧化剂。
参考
- Aladjadjiyan A.2002。磁场对玉米某些生物学特性影响的研究。中欧农业学报3:89-94。
- Alorainy, a . 2003。关于手机效应的最新研究。电磁场中的非电离辐射与我们的健康国际会议。分析生物化学72:248-254。
- Alscher酒店,R.G.Ertuk,N和希思,L.S.(2002)。在实验植物学plants.Journal控制氧化应激超氧化物歧化酶的作用。327(9):1331-1341。
- 阿佩尔,K. nadHirt,H.(2004)。活性氧:代谢,氧化胁迫和信号转导。植物生物学的年度审查。55(5):373-399。
- 亚迪。1949.甜菜叶绿体多酚氧化酶中铜酶的研究。植物生理学。24:1-15。
- Ayraapetyan,G. 2006. EMF波对大麦种子水合和萌发潜力的影响。电磁波和应用4:65-76。
- 贝茨,LS。, Walderen, RD. and Taere, ID。1973.水分胁迫研究中游离脯氨酸的快速测定。植物与土壤39:205-207。
- 弱磁场对植物的生物效应。空间研究进展34:1566-1574。
- Cakmak, T., Dumlupinar, R.和Erdal, S. 2009。不同磁场和渗透条件下小麦和豆苗萌发和早期生长的加速。Bioelectromagnetics。31:120 - 129。
- 陈志强,陈志强(2003)。抗氧化剂、氧化损伤和缺氧应激。《植物学。91(4): 179 - 194。
- 考德威尔,m.m.,罗伯雷切特,R.和弗林特,S.D.(1983)。内部过滤器:高等植物紫外驯化的前景。生理上的杆菌。58(8) 445 - 450。
- 张超,杨敏。(2002)。用两种互补比色法测定蜂胶中黄酮的含量。食品药品分析杂志。10(1) 178 - 182。
- 丁德萨,R.R.和Motowe,W.(1981).Drough耐受两种苔藓:与抗脂质过氧化酶防御的相关性。中华实验植物学。32(4):79-91。
- 弗林特,s.d.,乔丹,P.W.和考德威尔,M.M.(1985)。田间条件下增强duv - b辐射的植物保护反应:叶片光学特性和光合作用。光化学和光生物学杂志。41(4): 95 - 99。
- 张磊,高欧。(2008)。uv - b诱导拟南芥抗坏血酸缺失vtc1突变体的氧化应激和抗氧化防御系统响应。植物生理学杂志。22(4): 138 - 148。
- Gianopolitis, C.和Ries, S.K.(1977)。高等植物中超氧化物歧化酶的发生。植物生理学杂志。59(2): 309 - 314。
- 吉尔,S.S.和tuteja, N.(2010)。活性氧和抗氧化机制在作物抗非生物胁迫中的作用。植物生理学和生物化学。132(3): 909 - 930。
- 野兔,P.D.和Cress,W.a.(1997)。应激诱导的植物中的聚集在植物中的多样性原因。植物生长调控。21(2):79-103。
- Hosseinisarghein, S., Carapetian, J. and Khara, J.(2011)。本文讨论了紫外辐射对结构参数和超结构参数的影响。土耳其研究杂志。35(8): 69 - 77。
- IIao,学士(1997)。上植物学杂志tomato.Canadian的生长和光合作用紫外线-B-辐射和CO 2的影响。75(2):213-219。
- Jansen, M., gaba, V.和Greenberg, B.M.(1998)。高等植物与UV-B辐射:平衡损害、修复和积累。植物科学的趋势。3(1): 131 - 135。
- Lichtenthaler,香港(1987)。叶绿素和类胡萝卜素:光合生物膜的色素。nzymology的方法。148(4): 350 - 382。
- Nakano,Y.,Asada,K。(1981)。通过菠菜叶绿体中的抗坏血酸特异性过氧化物酶清除过氧化氢。植物细胞物理。22(1):867-880。
- 马乔,A.,宫崎骏,S.和Veronese P.(2002)。脯氨酸积累在胁迫诱导的生长减缓中发挥积极作用吗?植物杂志。31(5): 699 - 712。
- McDonald, S., Prenzler, P.D ., Autolovich, M.和Robards, K.(2001)。橄榄提取物的酚含量和抗氧化活性的食品化学。73(9): 73 - 84。
- Mittler, r(2002)。氧化应激,抗氧化剂和应激耐受性。植物科学的趋势。7(1): 405 - 410。
- Sakihama, Y., Cohen, M.F, Grace, S.C. and Yamaski, H>(2002)。植物酚类抗氧化和过氧化物活性:酚类诱导植物金属介导的氧化损伤。毒理学。177(9):67 - 80。
- 邵,宋,楚,L.Y, L.U.康,廖俊郎(2008)。高等植物细胞中初级抗氧化自由基清除和氧化还原信号通路。国际生物科学杂志。4(1): 8 - 14。
- Siripornadulsil, S., Traina, S., Vrma, D.P.和sayer, R.T.(2002)。脯氨酸介导的转基因微藻对有毒重金属耐受的分子机制植物细胞。14(7): 217 - 243。
- 司木露:(1995)。抗氧化系统和植物对环境的反应;在环境和植物代谢中。生物科学出版社,牛津,英国。111(2) 217 - 243。
- Tevini,M.,Braun,J.和Fieser,G.(1991)。Rye幼苗表皮层的生产功能对紫外线辐射。光化学和Pherobiology.verbruggen,N.,Hermans,C.(2008)。植物中的脯氨酸积累:审查。氨基酸。35(2):753-759。
- 姚,Y.,Xuana,Z.和Li,Y.(2006)。紫外线-B辐射对TartaryBucj小麦uder田间条件的作物生长,发育,产量和叶片色素浓度的影响。中国农艺学杂志。25(7):215-222。

这个作品是根据知识共享署名4.0国际许可.





